1. Барьерная - обеспечивает регулируемый, избирательный, пассивный и активный обмен веществ с окружающей средой.
Клеточные мембраны обладают избирательной проницаемостью : через них медленно диффундируют глюкоза, аминокислоты, жирные кислоты, глицерол и ионы, мембраны сами активно регулируют этот процесс - одни вещества пропускают, а другие нет.
2. Транспортная - через мембрану происходит транспорт веществ в клетку и из клетки. Транспорт через мембраны обеспечивает: доставку питательных веществ, удаление конечных продуктов обмена, секрецию различных веществ, создание ионных градиентов, поддержание в клетке соответствующего pH и ионной концентрации, которые нужны для работы клеточных ферментов.
Существует четыре основных механизма для поступления веществ в клетку или вывода их из клетки наружу:
а) Пассивный (диффузия, осмос) (не требует затрат энергии)
Диффузия
Распространение молекул или атомов одного вещества между молекулами или атомами другого, приводящее к самопроизвольному выравниванию их концентраций по всему занимаемому объёму. В некоторых ситуациях одно из веществ уже имеет выравненную концентрацию и говорят о диффузии одного вещества в другом. При этом перенос вещества происходит из области с высокой концентрацией в область с низкой концентрацией (вдоль вектора градиента концентрации (рис. 2.4).
Рис. 2.4. Схема процесса диффузии
Осмос
Процесс односторонней диффузии через полупроницаемую мембрану молекул растворителя в сторону бо́льшей концентрации растворённого вещества из объёма с меньшей концентрацией растворенного вещества (рис. 2.5).

Рис. 2.5. Схема процесса осмоса
б) Активный транспорт (требует затрат энергии)
Калий-натриевый насос (sodium-potassium pump) - механизм активного сопряженного трансмембранного транспорта ионов натрия (из клетки) и ионов калия (внутрь клетки), который обеспечивает концентрационный градиент и трансмембранную разность потенциалов. Последняя служит основой многих функций клеток и органов: секреции клеток желез, сокращения мышц, проведения нервных импульсов и др.(рис. 2.6).

Рис. 2.6. Схема работы калиево-натриевого насоса
На первой стадии фермент Na + /K + -АТФаза присоединяет с внутренней стороны мембраны три иона Na + . Эти ионы изменяют конформацию активного центра АТФ-азы. После этого фермент способен гидролизовать одну молекулу АТФ. Выделившаяся после гидролиза энергия расходуется на изменение конформации переносчика, благодаря чему три иона Na + и ион PO 4 3− (фосфат) оказываются на внешней стороне мембраны. Здесь ионы Na + отщепляются, а PO 4 3− замещается на два иона К + . После этого фермент возвращается в исходную конформацию, и ионы К + оказываются на внутренней стороне мембраны. Здесь ионы К + отщепляются, и переносчик вновь готов к работе.
В итоге во внеклеточной среде создается высокая концентрация ионов Na + , а внутри клетки - высокая концентрация K + . Эта разность концентраций используется в клетках при проведении нервного импульса.
в) Эндоцитоз (фагоцитоз, пиноцитоз)
Фагоцитоз (поедание клеткой) - процесс поглощения клеткой твёрдых объектов, таких как клетки эукариот, бактерии, вирусы, остатки мёртвых клеток и т. п. Вокруг поглощаемого объекта образуется большая внутриклеточная вакуоль (фагосома). Размер фагосом - от 250 нм и больше. Путем слияния фагосомы с первичной лизосомой образуется вторичная лизосома. В кислой среде гидролитические ферменты расщепляют макромолекулы, оказавшиеся во вторичной лизосоме. Продукты расщепления (аминокислоты, моносахариды и прочие полезные вещества) транспортируются затем через лизосомную мембрану в цитоплазму клетки. Фагоцитоз распространен очень широко. У высокоорганизованных животных и человека процесс фагоцитоза играет защитную роль. Фагоцитарная деятельность лейкоцитов и макрофагов имеет огромное значение в защите организма от попадающих в него патогенных микробов и других нежелательных частиц. Фагоцитоз впервые описал русский ученый И. И. Мечников(рис. 2.7)
Пиноцитоз (питьё клеткой) - процесс поглощения клеткой жидкой фазы из окружающей среды, содержащей растворимые вещества, включая крупные молекулы (белки, полисахариды и др.). При пиноцитозе от мембраны отшнуровываются внутрь клетки небольшие пузырьки - эндосомы. Они меньше фагосом (их размер до 150 нм) и обычно не содержат крупных частиц. После образования эндосомы к ней подходит первичная лизосома, и эти два мембранных пузырька сливаются. Образовавшаяся органелла носит название вторичной лизосомы. Процесс пиноцитоза постоянно осуществляют все эукариотические клетки (рис. 7)
Рецептор-опосредованный эндоцитоз - активный специфический процесс, при котором клеточная мембрана выпучивается внутрь клетки, формируя окаймлённые ямки. Внутриклеточная сторона окаймлённой ямки содержит набор адаптивных белков. Макромолекулы, связывающиеся со специфическими рецепторами на поверхности клетки, проходят внутрь со значительно большей скоростью, чем вещества, поступающие в клетки за счет пиноцитоза.
Рис. 2.7. Эндоцитоз
г) Экзоцитоз (отрицательный фагоцитоз и пиноцитоз)
Клеточный процесс, при котором внутриклеточные везикулы (мембранные пузырьки) сливаются с внешней клеточной мембраной. При экзоцитозе содержимое секреторных везикул (экзоцитозных пузырьков) выделяется наружу, а их мембрана сливается с клеточной мембраной. Практически все макромолекулярные соединения (белки, пептидные гормоны и др.) выделяются из клетки этим способом (рис. 2.8)
Рис. 2.8. Схема экзоцитоза
3. Генерация и проведение биопотенциалов - с помощью мембраны в клетке поддерживается постоянная концентрация ионов: концентрация иона К+ внутри клетки значительно выше, чем снаружи, а концентрация Na+ значительно ниже, что очень важно, так как это обеспечивает поддержание разности потенциалов на мембране и генерацию нервного импульса.
4. Механическая - обеспечивает автономность клетки, ее внутриклеточных структур, также соединение с другими клетками (в тканях).
5. Энергетическая - при фотосинтезе в хлоропластах и клеточном дыхании в митохондриях в их мембранах действуют системы переноса энергии, в которых также участвуют белки;
6. Рецепторная - некоторые белки, находящиеся в мембране, являются рецепторами (молекулами, при помощи которых клетка воспринимает те или иные сигналы).
7. Ферментативная - мембранные белки нередко являются ферментами. Например, плазматические мембраны эпителиальных клеток кишечника содержат пищеварительные ферменты.
8. Матричная - обеспечивает определенное взаиморасположение и ориентацию мембранных белков, их оптимальное взаимодействие;
9. Маркировка клетки - на мембране есть антигены, действующие как маркеры - «ярлыки», позволяющие опознать клетку. Это гликопротеины (то есть белки с присоединенными к ним разветвленными олигосахаридными боковыми цепями), играющие роль «антенн». С помощью маркеров клетки могут распознавать другие клетки и действовать согласованно с ними, например, при формировании органов и тканей. Это же позволяет иммунной системе распознавать чужеродные антигены.
Клеточные включения
К клеточным включениям относятся углеводы, жиры и белки. Все эти вещества накапливаются в цитоплазме клетки в виде капель и зерен различной величины и формы. Они периодически синтезируются в клетке и используются в процессе обмена веществ.
Цитоплазма
Это часть живой клетки (протопласта) без плазматической мембраны и ядра. В состав цитоплазмы входят: цитоплазматический матрикс, цитоскелет, органоиды и включения (иногда включения и содержимое вакуолей к живому веществу цитоплазмы не относят). Отграниченная от внешней среды плазматической мембраной, цитоплазма представляет собой внутреннюю полужидкую среду клеток. В цитоплазму эукариотических клеток располагаются ядро и различные органоиды. В ней сосредоточены и разнообразные включения - продукты клеточной деятельности, вакуоли, а также мельчайшие трубочки и нити, образующие скелет клетки. В составе основного вещества цитоплазмы преобладают белки.
Функции цитоплазмы
1) в ней протекают основные процессы обмена веществ.
2) объединяет в одно целое ядро и все органоиды, обеспечивает их взаимодействие.
3) подвижность, раздражимость, метаболизм и размножение.
Подвижность проявляется в различных формах:
Внутриклеточное движение цитоплазмы клетки.
Амебовидное движение. Эта форма движения выражается в образовании цитоплазмой псевдоподий в сторону того или иного раздражителя или от него. Эта форма движения присуща амебе, лейкоцитам крови, а также некоторым тканевым клеткам.
Мерцательное движение. Проявляется в виде биений крошечных протоплазматических выростов - ресничек и жгутиков (инфузории, клетки эпителия многоклеточных животных, спермии и др.).
Сократительное движение. Обеспечивается благодаря присутствию в цитоплазме специального органоида миофибрилл, укорочение или удлинение которого способствуют сокращению и расслаблению клетки. Способность к сокращению наиболее развита у мышечных клеток.
Раздражимость выражается в способности клеток реагировать на раздражение изменением обмена веществ и энергии.
Цитоскелет
Одной из отличительных особенностей эукариотической клетки является наличие в ее цитоплазме скелетных образований в виде микротрубочек и пучков белковых волокон. Элементы цитоскелета, тесно связанные с наружной цитоплазматической мембраной и ядерной оболочкой, образуют сложные переплетения в цитоплазме.
Цитоскелет образован микротрубочками, микрофиламентами и микротрабекулярной системой. Цитоскелет определяет форму клетки, участвует в движениях клетки, в делении и перемещениях самой клетки, во внутриклеточном транспорте органоидов.
Микротрубочки содержатся во всех эукариотических клетках и представляют собой полые неразветвленные цилиндры, диаметр которых не превышает 30 нм, а толщина стенки - 5 нм. В длину они могут достигать нескольких микрометров. Легко распадаются и собираются вновь. Стенка микротрубочек в основном построена из спирально уложенных субъединиц белка тубулина (рис. 2.09)
Функции микротрубочек :
1) выполняют опорную функцию;
2) образуют веретено деления; обеспечивают расхождение хромосом к полюсам клетки; отвечают за перемещение клеточных органелл;
3) принимают участие во внутриклеточном транспорте, секреции, формировании клеточной стенки;
4) являются структурным компонентом ресничек, жгутиков, базальных телец и центриолей.
Микрофиламенты представлены нитями диаметром 6 нм, состоящими из белка актина, близкого к актину мышц. Актин составляет 10-15% общего количества белка клетки. В большинстве животных клеток образуется густая сеть из актиновых филаментов и связанных с ними белков под самой плазматической мембраной.
Помимо актина, в клетке обнаруживаются и нити миозина. Однако количество их значительно меньше. Благодаря взаимодействию актина и миозина происходит сокращение мышц. Микрофиламенты связаны с движением всей клетки либо ее отдельных структур внутри нее. В некоторых случаях движение обеспечивается только актиновыми филаментами, в других - актином вместе с миозином.
Функции микрофиламентов
1) механическая прочность
2) позволяет клетке изменять свою форму и двигаться.

Рис. 2.09. Цитоскелет
Органоиды (или органеллы)
Делятся на немембранные, одномембранные и двумембранные .
К немембранным органоидам эукариотической клетки относятся органоиды, не имеющие собственной замкнутой мембраны, а именно: рибосомы и органоиды, построенные на основе тубулиновых микротрубочек – клеточный центр (центриоли) и органоиды движения (жгутики и реснички). В клетках большинства одноклеточных организмов и подавляющего большинства высших (наземных) растений центриоли отсутствуют.
К одномембранным органоидам относятся: эндоплазматическая сеть, аппарат Гольджи, лизосомы, пероксисомы, сферосомы, вакуоли и некоторые другие. Все одномембранные органоиды связаны между собой в единую систему клетки. В растительных клетках имеются особенные лизосомы, в животных клетках имеются особенные вакуоли: пищеварительные, выделительные, сократительные, фагоцитарные, аутофагоцитарные и др.
К двумембранным органоидам относятся митохондрии и пластиды.
Немембранные органоиды
А) Рибосомы – органоиды, встречающиеся в клетках всех организмов. Это мелкие органеллы, представленные глобулярными частицами диаметром порядка 20 нм. Рибосомы состоят из двух субъединиц неравного размера - большой и малой. В состав рибосом входят белки и рибосомальные РНК (рРНК). Различают два основных типа рибосом: эукариотические (80S) и прокариотические (70S).
В зависимости от локализации в клетке, различают свободные рибосомы, находящиеся в цитоплазме, синтезирующие белки и прикрепленные рибосомы - рибосомы, связанные большими субъединицами с наружной поверхностью мембран ЭПР, синтезирующие белки, которые поступают в комплекс Гольджи, а затем секретируются клеткой. Во время биосинтеза белка рибосомы могут объединяться в комплексы - полирибосомы (полисомы).
Рибосомы эукариот образуются в ядрышке. Сначала на ядрышковой ДНК синтезируются рРНК, которые затем покрываются поступающими из цитоплазмы рибосомальными белками, расщепляются до нужных размеров и формируют субъединицы рибосом. Полностью сформированных рибосом в ядре нет. Объединение субъединиц в целую рибосому происходит в цитоплазме, как правило, во время биосинтеза белка.
Рибосомы обнаружены в клетках всех организмов. Каждая состоит из двух частиц, малой и большой. В состав рибосом входят белки и РНК.
Функции
синтез белка.
Синтезированные белки сначала накапливаются в каналах и полостях эндоплазматической сети, а затем транспортируются к органоидам и участкам клетки. ЭПС и рибосомы, расположенные на ее мембранах, представляют собой единый аппарат биосинтеза и транспортировки белков (рис. 2.10-2.11).
Рис. 2.10. Строение рибосомы

Рис. 2.11. Строение рибосом
В) Клеточный центр (центриоли)
Центриоль представляет собой цилиндр (длиной 0,3 мкм и диаметром 0,1 мкм), стенка которого образована девятью группами из трех слившихся микротрубочек (9 триплетов), соединенных между собой через определенные интервалы поперечными сшивками. Часто центриоли объединены в пары, где они расположены под прямым углом друг к другу. Если центриоль лежит в основании реснички или жгутика, то ее называют базальным тельцем.
Почти во всех животных клетках имеется пара центриолей, являющихся срединным элементом клеточного центра.
Перед делением центриоли расходятся к противоположным полюсам и возле каждой из них возникает дочерняя центриоль. От центриолей, расположенных на разных полюсах клетки, образуются микротрубочки, растущие навстречу друг другу.
Функции
1) формируют митотическое веретено, способствующее равномерному распределению генетического материала между дочерними клетками,
2) являются центром организации цитоскелета. Часть нитей веретена прикрепляется к хромосомам.
Центриоли относятся к самовоспроизводящимся органоидам цитоплазмы. Они возникают в результате дупликации уже имеющихся. Это происходит при расхождении центриолей. Незрелая центриоль содержит 9 одиночных микротрубочек; по-видимому, каждая микротрубочка является матрицей при сборке триплетов, характерных для зрелой центриоли(рис. 2.12).
Цетриоли имеются в клетках низших растений (водоросли).

Рис. 2.12. Центриоли клеточного центра
Одномембранные органоиды
Г) Эндоплазматическая сеть (ЭПС)
Вся внутренняя зона цитоплазмы заполнена многочисленными мелкими каналами и полостями, стенки которых представляют собой мембраны, сходные по своей структуре с плазматической мембраной. Эти каналы ветвятся, соединяются друг с другом и образуют сеть, получившую название эндоплазматической сети. Эндоплазматическая сеть неоднородна по своему строению. Известны два ее типа - гранулярная и гладкая .
На мембранах каналов и полостей гранулярной сети располагается множество мелких округлых телец - рибосом , которые придают мембранам шероховатый вид. Мембраны гладкой эндоплазматической сети не несут рибосом на своей поверхности. ЭПС выполняет много разнообразных функций.
Функции
Основная функция гранулярной эндоплазматической сети - участие в синтезе белка, который осуществляется в рибосомах. На мембранах гладкой эндоплазматической сети происходит синтез липидов и углеводов. Все эти продукты синтеза накапливаются в каналах и полостях, а затем транспортируются к различным органоидам клетки, где потребляются или накапливаются в цитоплазме в качестве клеточных включений. ЭПС связывает между собой основные органоиды клетки(рис. 2.13).
Рис. 2.13. Строение эндоплазматической сети (ЭПС) или ретикулума
Д) Аппарат Гольджи
Строение этого органоида сходно в клетках растительных и животных организмов, несмотря на разнообразие его формы. Выполняет много важных функций.
Одномембранный органоид. Представляет собой стопки уплощенных «цистерн» с расширенными краями, с которыми связана система мелких одномембранных пузырьков (пузырьки Гольджи). Пузырьки Гольджи в основном сконцентрированы на стороне, примыкающей к ЭПС, и по периферии стопок. Полагают, что они переносят в аппарат Гольджи белки и липиды, молекулы которых, передвигаясь из цистерны в цистерну, подвергаются химической модификации.
Все эти вещества сначала накапливаются, химически усложняются, а затем в виде крупных и мелких пузырьков поступают в цитоплазму и либо используются в самой клетке в процессе ее жизнедеятельности, либо выводятся из нее и используются в организме (рис. 2.14-2.15).
Рис. 2.14. Строение аппарата Гольджи
Функции :
Модификация и накопление белков, липидов, углеводов;
Упаковка в мембранные пузырьки (везикулы) поступивших органических веществ;
Место образования лизосом;
Секреторная функция, поэтому аппарат Гольджи хорошо развит в секреторных клетках.

Рис. 2.15. Комплекс Гольджи
Е) Лизосомы
Представляют собой небольшие округлые тельца. Внутри лизосомы находятся ферменты, расщепляющие белки, жиры, углеводы, нуклеиновые кислоты. К пищевой частице, поступившей в цитоплазму, подходят лизосомы, сливаются с ней, и образуется одна пищеварительная вакуоль, внутри которой находится пищевая частица, окруженная ферментами лизосом.
Ферменты лизосом синтезируются на шероховатой ЭПС, перемещаются в аппарат Гольджи, где происходит их модификация и упаковка в мембранные пузырьки лизосом. Лизосома может содержать от 20 до 60 различных видов гидролитических ферментов. Расщепление веществ с помощью ферментов называют лизисом .
Различают первичные и вторичные лизосомы. Первичными называются лизосомы, отпочковавшиеся от аппарата Гольджи.
Вторичными называются лизосомы, образовавшиеся в результате слияния первичных лизосом с эндоцитозными вакуолями. В этом случае в них происходит переваривание веществ, поступивших в клетку путем фагоцитоза или пиноцитоза, поэтому их можно назвать пищеварительными вакуолями.
Функции лизосом:
1) переваривание захваченных клеткой при эндоцитозе веществ или частиц (бактерий, других клеток),
2) аутофагия - уничтожение ненужных клетке структур, например, во время замены старых органоидов новыми, или переваривание белков и других веществ, произведенных внутри самой клетки,
3) автолиз - самопереваривание клетки, приводящее к ее гибели (иногда этот процесс не является патологическим, а сопровождает развитие организма или дифференцировку некоторых специализированных клеток) (рис. 2.16-2.17).
Пример: При превращении головастика в лягушку, лизосомы, находящиеся в клетках хвоста, переваривают его: хвост исчезает, а образовавшиеся во время этого процесса вещества всасываются и используются другими клетками тела.
Рис. 2.16. Образование лизосом

Рис. 2.17. Функционирование лизосом
Ж) Пероксисомы
Органоиды, сходные по строению с лизосомами, пузырьки с диаметром до 1,5 мкм с однородным матриксом, содержащим около 50 ферментов.
Каталаза вызывает распад перекиси водорода 2Н 2 О 2 → 2Н 2 О + О 2 и предотвращает перекисное окисление липидов
Образуются пероксисомы отпочковываваясь от ранее существующих, т.е. относятся к самовоспроизводящимся органоидам, несмотря на то, что не содержат ДНК. Растут благодаря поступлению в них ферментов, ферменты пероксисом образуются на шероховатой ЭПС и в гиалоплазме (рис. 2.18) .
Рис. 2.18. Пероксисома (в центре кристаллический нуклеоид)
З) Вакуоли
Одномембранные органоиды. Вакуоли представляют собой «емкости», заполненные водными растворами органических и неорганических веществ. В образовании вакуолей принимают участие ЭПС и аппарат Гольджи.
Молодые растительные клетки содержат много мелких вакуолей, которые затем по мере роста и дифференцировки клетки сливаются друг с другом и образуют одну большую центральную вакуоль.
Центральная вакуоль может занимать до 95% объема зрелой клетки, ядро и органоиды оттесняются при этом к клеточной оболочке. Мембрана, ограничивающая растительную вакуоль, называется тонопластом .
Жидкость, заполняющая растительную вакуоль, называется клеточным соком. В состав клеточного сока входят водорастворимые органические и неорганические соли, моносахариды, дисахариды, аминокислоты, конечные или токсические продукты обмена веществ (гликозиды, алкалоиды), некоторые пигменты (антоцианы).
Из органических веществ чаще запасаются сахара и белки. Сахара – чаще в виде растворов, белки поступают в виде пузырьков ЭПР и аппарата Гольджи, после чего вакуоли обезвоживаются, превращаясь в алейроновые зерна.
В животных клетках имеются мелкие пищеварительные и автофагические вакуоли, относящиеся к группе вторичных лизосом и содержащие гидролитические ферменты. У одноклеточных животных есть еще сократительные вакуоли, выполняющие функцию осморегуляции и выделения.
Функции
У растений
1) накопление жидкости и поддержание тургора,
2) накопление запасных питательных веществ и минеральных солей,
3) окрашивание цветов и плодов и привлечение тем самым опылителей и распространителей плодов и семян.
У животных:
4) пищеварительные вакуоли – разрушают органические макромолекулы;
5) сократительные вакуоли регулируют осмотическое давление клетки и выводят ненужные вещества из клетки
6) фагоцитарные вакуоли образуются при фагоцитозе иммунными клетками антигенов
7) аутофагоцитарные вакуоли образуются при фагоцитозе иммунными клетками собственных тканей
Двумембранные органоиды (митохондрии и пластиды)
Эти органоиды являются полуавтономными, поскольку обладают собственной ДНК и собственным белоксинтезирующим аппаратом. Митохондрии имеются практически во всех эукариотических клетках. Пластиды имеются только в растительных клетках.
И) Митохондрии
Это органеллы энергообеспечения метаболических процессов в клетке. В гиалоплазме митохондрии распределены обычно диффузно, однако в специализированных клетках сосредоточены в тех участках, где имеется наибольшая потребность в энергии. Например, в мышечных клетках большие количества митохондрий сосредоточены вдоль сократительных фибрилл, вдоль жгутика сперматозоида, в эпителии почечных канальцев, в области синапсов и т. д. Такое расположение митохондрий обеспечивает меньшие потери АТФ во время ее диффузии.
Наружная мембрана отделяет митохондрию от цитоплазмы, замкнута сама на себя и не образует впячиваний. Внутренняя мембрана ограничивает внутреннее содержимое митохондрий – матрикс. Характерная особенность – образование многочисленных впячиваний – крист, за счет чего площадь внутренних мембран увеличивается. Количество и степень развития крист зависит от функциональной активности ткани. Митохондрии имеют собственный генетический материал (рис. 2.19).
ДНК митохондрий – это замкнутая кольцевая двуспиральная молекула, в клетках человека имеет размер 16569 нуклеотидных пар, это приблизительно в 105 раз меньше ДНК, локализованной в ядре. Митохондрии обладают собственной белоксинтезирующей системой, количество же транслируемых с митохондриальной мРНК белков ограничено. Митохондриальные ДНК не могут кодировать все митохондриальные белки. Большая часть белков митохондрий находится под генетическим контролем ядра.

Рис. 2.19. Строение митохондрий
Функции митохондрий
1) образование АТФ
2) синтез белка
3) участие в специфических синтезах, например, синтез стероидных гормонов (надпочечники)
4) отработавшие митохондрии могут накапливать и продукты экскреции, вредные вещества, т.е. способны брать на себя функции других органелл клетки
К) Пластиды
Пластиды –органеллы, характерные только для растений.
Различают три типа пластид:
1) хлоропласты (пластиды зеленого цвета);
2) хромопласты (пластиды желтого, оранжевого или красного цвета)
3) лейкопласты (бесцветные пластиды).
Обычно в клетке встречаются пластиды только одного типа.
Хлоропласты
Эти органоиды содержатся в клетках листьев и других зеленых органов растений, а также у разнообразных водорослей. У высших растений в одной клетке обычно бывает несколько десятков хлоропластов. Зеленый цвет хлоропластов зависит от содержания в них пигмента хлорофилла.
Хлоропласт - основной органоид клеток растений, в котором происходит фотосинтез, т. е. образование органических веществ (углеводов) из неорганических (СО 2 и Н 2 О) при использовании энергии солнечного света. По строению хлоропласты сходны с митохондриями.
Хлоропласты имеют сложное строение. От гиалоплазмы они отграничены двумя мембранами – наружной и внутренней. Внутреннее содержимое называется строма . Внутренняя мембрана формирует внутри хлоропласта сложную, строго упорядоченную систему мембран, имеющих форму плоских пузырьков, называемых тилакоидами .
Тилакоиды собраны в стопки - граны , напоминающие столбики монет. Граны связаны между собой тилакоидами стромы, проходящими через них насквозь вдоль пластиды (рис. 2.20-2.22). Хлорофилл и хлоропласты образуются только на свету.
Рис. 2.20. Хлоропласты под световым микроскопом
Рис. 2.21. Строение хлоропласта под электронным микроскопом

Рис. 2.22. Схематичное строение хлоропластов
Функции
1) фотосинтез (образование органических веществ из неорганических веществ за счет энергии света). Центральная роль в этом процессе принадлежит хлорофиллу. Он поглощает энергию света и направляет ее на осуществление реакций фотосинтеза. В хлоропластах, как и в митохондриях, происходит синтез АТФ.
2) участвуют в синтезе аминокислот и жирных кислот,
3) служат хранилищем временных запасов крахмала.
Лейкопласты - мелкие бесцветные пластиды, которые встречаются в клетках органов, скрытых от солнечного света (корни, корневища, клубни, семена). Строение их сходно со строением хлоропластов (рис. 2.23).
Однако, в отличие от хлоропластов, у лейкопластов слабо развита внутренняя мембранная система, т.к. они участвуют в синтезе и накоплении запасных питательных веществ - крахмала, белков и липидов. На свету лейкопласты могут превращаться в хлоропласты.
Рис. 2.23. Строение лейкопласта
Хромопласты - пластиды оранжевого, красного и желтого цвета, который обусловлен пигментами, относящимися к группе каротиноидов. Хромопласты встречаются в клетках лепестков многих растений, зрелых плодов, редко - корнеплодов, а также в осенних листьях. Внутренняя мембранная система в хромопластах, как правило, отсутствует (рис. 24) .
Рис. 2.24. Строение хромопласта
Значение хромопластов до конца еще не выяснено. Большинство из них представляют собой стареющие пластиды. Они, как правило, развиваются из хлоропластов, при этом в пластидах разрушаются хлорофилл и внутренняя мембранная структура, и накапливаются каротиноиды. Это происходит при созревании плодов и пожелтении листьев осенью. Биологическое значение хромопластов состоит в том, что они обусловливают яркую окраску цветков и плодов, привлекающую насекомых для перекрестного опыления и других животных для распространения плодов. В хромопласты могут превращаться и лейкопласты.
Функции пластид
Синтез в хлорофилле органических веществ из простых неорганических соединений: углекислого газа и воды в присутствии квантов солнечного света – фотосинтез, синтез АТФ в световую фазу фотосинтеза
Синтез белков на рибосомах (между внутренними мембранами хлоропласта содержатся ДНК, РНК и рибосомы, следовательно, в хлоропластах, так же как и в митохондриях, происходит синтез белка, необходимого для деятельности этих органоидов).
Присутствие хромопластов объясняется желтая, оранжевая и красная окраска венчиков цветков, плодов, осенних листьев.
Лейкопласты содержат запасающие вещества (в стеблях, корнях, клубнях).
Хлоропласты, хромопласты и лейкопласты способны клетка взаимному переходу. Так при созревании плодов или изменении окраски листьев осенью хлоропласты превращаются в хромопласты, а лейкопласты могут превращаться в хлоропласты, например, при приобретении зеленого цвета клубней картофеля.
В эволюционном смысле первичным, исходным типом пластид являются хлоропласты, из которых произошли пластиды остальных двух типов. Пластиды имеют много общих черт с митохондриями, отличающих их от других компонентов цитоплазмы. Это, прежде всего, оболочка из двух мембран и относительная генетическая автономность, обусловленная наличием собственных рибосом и ДНК. Такое своеобразие органелл легло в основу представления, что предшественниками пластид и митохондрий были бактерии, которые в процессе эволюции оказались встроенными в эукариотическую клетку и постепенно превратились в хлоропласты и митохондрии (рис. 2.25).

Рис. 2.25. Образование митохондрий и хлоропластов по теории симбиогенеза
Плазматическая мембрана выполняет ряд важнейших функций:
1) Барьерная. Барьерная функция плазматической мембраны заключается в ограничении свободной диффузии веществ из клетки в клетку, предотвращении утечки водорастворимого содержимого клетки. Но поскольку клетка должна получать необходимые питательные вещества, выделять конечные продукты метаболизма, регулировать внутриклеточные концентрации ионов, то в ней образовались специальные механизмы переноса веществ через клеточную мембрану.
2) Транспортная. К транспортной функции относится обеспечение поступления и выведения различных веществ в клетку и из клетки. Важное свойство мембраны - избирательная проницаемость , или полупроницаемость. Она легко пропускает воду и водорастворимые газы и отталкивает полярные молекулы, такие как глюкоза или аминокислоты.
Существует несколько механизмов транспорта веществ через мембрану:
пассивный транспорт;
активный транспорт;
транспорт в мембранной упаковке.
Пассивный транспорт. Диффузия - это движение частиц среды, приводящее к переносу вещества из зоны, где его концентрация высока в зону с низкой концентрацией. При диффузионном транспорте мембрана функционирует как осмотический барьер. Скорость диффузии зависит от величины молекул и их относительной растворимости в жирах. Чем меньше размеры молекул и чем более они жирорастворимы (липофильны), тем быстрее произойдет их перемещение через липидный бислой. Диффузия может быть нейтральной (перенос незаряженных молекул) и облегченной (с помощью специальных белков переносчиков). Скорость облегченной диффузии выше, чем нейтральной. Максимальной проникающей способностью обладает вода, так как ее молекулы малы и незаряжены. Диффузия воды через клеточную мембрану называется осмосом. Предполагается, что в клеточной мембране для проникновения воды и некоторых ионов существуют специальные "поры". Число их невелико, а диаметр составляет около 0,3-0,8 нм. Наиболее быстро диффундируют через мембрану легко растворимые в липидном бислое молекулы, например О, и незаряженные полярные молекулы небольшого диаметра (СО, мочевина).
Перенос полярных молекул (сахаров, аминокислот), осуществляемый с помощью специальных мембранных транспортных белков называется облегченной диффузией. Такие белки обнаружены во всех типах биологических мембран, и каждый конкретный белок предназначен для переноса молекул определенного класса. Транспортные белки являются трансмембранными, их полипептидная цепь пересекает липидный бислой несколько раз, формируя в нем сквозные проходы. Это обеспечивает перенос специфических веществ через мембрану без непосредственного контакта с ней. Существует два основных класса транспортных белков: белки-переносчики (транспортеры) и каналообразующие белки (белки-каналы). Белки-переносчики переносят молекулы через мембрану, предварительно изменяя их конфигурацию. Каналообразующие белки формируют в мембране заполненные водой поры. Когда поры открыты, молекулы специфических веществ (обычно неорганические ионы подходящего размера и заряда) проходят сквозь них. Если молекула транспортируемого вещества не имеет заряда, то направление транспорта определяется градиентом концентрации. Если молекула заряжена, то на ее транспорт, кроме градиента концентрации, влияет и электрический заряд мембраны (мембранный потенциал). Внутренняя сторона плазмалеммы обычно заряжена отрицательно по отношению к наружной. Мембранный потенциал облегчает проникновение в клетку положительно заряженных ионов и препятствует прохождению ионов заряженных отрицательно.
Активный транспорт. Активным транспортом называется перенос веществ против электрохимического градиента. Он всегда осуществляется белками-транспортерами и тесно связан с источником энергии. В белках-переносчиках имеются участки связывания с транспортируемым веществом. Чем больше таких участков связывается с веществом, тем выше скорость транспорта. Селективный перенос одного вещества называется унипортом. Перенос нескольких веществ осуществляют котранспортные системы. Если перенос идет в одном направлении - это симпорт, если в противоположных - антипорт. Так, например, глюкоза из внеклеточной жидкости в клетку переносится унипортно. Перенос же глюкозы и Na 4 из полости кишечника или канальцев почек соответственно в клетки кишечника или кровь осуществляется симпортно, а перенос С1~ и НСО" антипортно. Предполагается, что при переносе возникают обратимые конформационные изменения в транспортере, что и позволяет премещать соединенные с ним вещества.
Примером белка-переносчика, использующего для транспорта веществ энергию выделившуюся при гидролизе АТФ, является Na + -К + насос, обнаруженный в плазматической мембране всех клеток. Na + -K насос работает по принципу антипорта, перекачивая Na" из клетки и К т внутрь клетки против их электрохимических градиентов. Градиент Na + создает осмотическое давление, поддерживает клеточный объем и обеспечивает транспорт сахаров и аминокислот. На работу этого насоса тратится треть всей энергии необходимой для жизнедеятельности клеток. При изучении механизма действия Na + -K + насоса было установлено, что он является ферментом АТФазой и трансмембранным интегральным белком. В присутствии Na + и АТФ под действием АТФа-зы от АТФ отделяется концевой фосфат и присоединяется к остатку аспарагиновой кислоты на молекуле АТФазы. Молекула АТФазы фосфорилируется, изменяет свою конфигурацию и Na + выводится из клетки. Вслед за выведением Na из клетки всегда происходит транспорт К" в клетку. Для этого от АТФазы в присутствии К отщепляется ранее присоединенный фосфат. Фермент дефосфорилируется, восстанавливает свою конфигурацию и К 1 "закачивается" в клетку.
АТФаза образована двумя субъединицами, большой и малой. Большая субъединица состоит из тысячи аминокислотных остатков, пересекающих бислой несколько раз. Она обладает каталитической активностью и способна обратимо фосфорилироваться и дефосфорилироваться. Большая субъединица на цитоплазматической стороне имеет участки для связывания Na + и АТФ, а на внешней стороне -участки для связывания К + и уабаина. Малая субъединица является гликопротеином и функция его пока не известна.
Na + -K насос обладает электрогенным эффектом. Он удаляет три положительно заряженных иона Na f из клетки и вносит в нее два иона К В результате через мембрану течет ток, образующий электрический потенциал с отрицательным значением во внутренней части клетки по отношению к ее наружной поверхности. Na"-K + насос регулирует клеточный объем, контролирует концентрацию веществ внутри клетки, поддерживает осмотическое давление, участвует в создании мембранного потенциала.
Транспорт в мембранной упаковке. Перенос через мембрану макромолекул (белков, нуклеиновых кислот, полисахаридов, липопротеидов) и других частиц осуществляется посредством последовательного образования и слияния окруженных мембраной пузырьков (везикул). Процесс везикулярного транспорта проходит в две стадии. Вначале мембрана пузырька и плазмалемма слипаются, а затем сливаются. Для протекания 2 стадии необходимо чтобы молекулы воды были вытеснены взаимодействующими липидными бислоями, которые сближаются до расстояния 1-5 нм. Считается, что данный процесс активизируют специальные белки слияния (они выделены пока только у вирусов). Везикулярный транспорт имеет важную особенность - поглощенные или секретируемые макромолекулы, находящиеся в пузырьках, обычно не смешиваются с другими макромолекулами или органеллами клетки. Пузырьки могут сливаться со специфическими мембранами, что и обеспечивает обмен макромолекулами между внеклеточным пространством и содержимым клетки. Аналогично происходит перенос макромолекул из одного компартмента клетки в другой.
Транспорт макромолекул и частиц в клетку называется эндоцитозом. При этом транспортируемые вещества обволакиваются частью плазматической мембраны, образуется пузырек (вакуоль), который перемещается внутрь клетки. В зависимости от размера образующихся пузырьков различают два вида эндоцитоза - пиноцитоз и фагоцитоз.
Пиноцитоз обеспечивает поглощение жидкости и растворенных веществ в виде небольших пузырьков (d=150 нм). Фагоцитоз - это поглощение больших частиц, микроорганизов или обломков органелл, клеток. При этом образуются крупные пузырьки, фагосомы или вакуоли (d-250 нм и более). У простейших фагоцитарная функция - форма питания. У млекопитающих фагоцитарная функция осуществляется макрофагами и нейтрофилами, защищающими организм от инфекции путем поглощения вторгшихся микробов. Макрофаги участвуют также в утилизации старых или поврежденных клеток и их обломков (в организме человека макрофаги ежедневно поглощают более 100 старых эритроцитов). Фагоцитоз начинается только тогда, когда поглощаемая частица свяжется с поверхностью фагоцита и активирует специализированные рецепторные клетки. Связывание частиц со специфическими рецепторами мембраны вызывает образование псевдоподии, которые обволакивают частицу и, сливаясь краями, образуют пузырек -фагосому. Образование фагосомы и собственно фагоцитоз происходит лишь в том случае, если в процессе обволакивания частица постоянно контактирует с рецепторами плазмалеммы, как бы "застегивая молнию".
Значительная часть материала, поглощенного клеткой путем эндоцитоза, заканчивает свой путь в лизосомах. Большие частицы включаются в фагосомы, которые затем сливаются с лизосомами и образуют фаголизосомы. Жидкость и макромолекулы, поглощенные при пиноцитозе, первоначально переносятся в эндосомы, которые также сливаются с лизосомами, образуя эндолизосомы. Присутствующие в лизосомах разнообразные гидролитические ферменты быстро разрушают макромолекулы. Продукты гидролиза (аминокислоты, сахара, нуклеотиды) транспортируются из лизосом в цитозоль, где используются клеткой. Большинство мембранных компонентов эндоцитозных пузырьков из фагосом и эндосом возвращаются с помощью экзоцитоза к плазматической мембране и там повторно утилизируются. Основным биологическим значением эндоцитоза является получение строительных блоков за счет внутриклеточного переваривания макромолекул в лизосомах.
Поглощение веществ в эукариотических клетках начинается в специализированных областях плазматической мембраны, так называемых окаймленных ямках. На электронных микрофотографиях ямки выглядят как впячивания плазматической мембраны, цитоплазматическая сторона которых покрыта волокнистым слоем. Слой как бы окаймляет небольшие ямки плазмалеммы. Ямки занимают около 2% общей поверхности клеточной мебраны эукариотов. В течении минуты ямки растут, все глубже впячиваются, втягиваются в клетку и затем, сужаясь у основания, отщепляются, образуя окаймленные пузырьки. Установлено, что из плазматической мембраны фибробластов в течении одной минуты отщепляется примерно четвертая часть мембраны в виде окаймленных пузырьков. Пузырьки быстро теряют свою кайму и приобретают способность сливаться с лизосомой.
Эндоцитоз может быть неспецифическим (конститутивным) и специфическим (рецепторным). При неспецифическом эндоцитозе клетка захватывает и поглощает совершенно чуждые ей вещества, например, частицы сажи, красители. Вначале происходит осаждение частиц на гликокаликсе плазмалеммы. Особенно хорошо осаждаются (адсорбируются) положительно заряженные группы белков, так как гликокаликс несет отрицательный заряд. Затем изменяется морфология клеточной мембраны. Она может либо погружаться, образуя впячивания (инвагинации), либо, наоборот, формировать выросты, которые как бы складываются, отделяя небольшие объемы жидкой среды. Образование инвагинаций более характерно для клеток кишечного эпителия, амеб, а выростов - для фагоцитов и фибробластов. Заблокировать эти процессы можно ингибиторами дыхания. Образовавшиеся пузырьки - первичные эндосомы, могут сливаться между собой, увеличиваясь в размере. В дальнейшем они соединяются с лизосомами, превращаясь в эндолизосому - пищеварительную вакуоль. Интенсивность жидкофазного неспецифического пиноцитоза довольно высока. Макрофаги образуют до 125, а клетки эпителия тонкого кишечника до тысячи пиносом в минуту. Обилие пиносом приводит к тому, что плазмалемма быстро тратится на образование множества мелких вакуолей. Восстановление мембраны идет довольно быстро при рециклизации в процессе экзоцитоза за счет возвращения вакуолей и их встраивания в плазмалемму. У макрофагов вся плазматическая мембрана замещается за 30 минут, а у фибробластов за 2 часа.
Более эффективным способом поглощения из внеклеточной жидкости специфических макромолекул является специфический эндоцитоз (опосредуемый рецепторами). Макромолекулы при этом связываются с комплементарными рецепторами на поверхности клетки, накапливаются в окаймленной ямке, и затем, образуя эндосому, погружаются в цитозоль. Рецепторный эндоцитоз обеспечивает накопление специфических макромолекул у своего рецептора. Молекулы, которые связываются на поверхности плазмалеммы с рецептором, называются лигандами. При помощи рецепторного эндоцитоза во многих животных клетках идет поглощение холестерина из внеклеточной среды.
Плазмолемма принимает участие в выведении веществ из клетки (экзоцитоз). В этом случае вакуоли подходят к плазмолемме. В местах контактов плазмолемма и мембрана вакуоли сливаются и содержимое вакуоли поступает в окружающую среду. У некоторых простейших места на клеточной мембране для экзоцитоза заранее предопределены. Так, в плазматической мембране некоторых ресничных инфузорий есть определенные участки с правильным расположением крупных глобул интегральных белков. У мукоцист и трихоцист инфузорий полностью готовых к секреции, на верхней части плазмалеммы имеется венчик из глобул интегральных белков. Этими участками мембраны мукоцист и трихоцист соприкасаются с поверхностью клетки. Своеобразный экзоцитоз наблюдается в нейтрофилах. Они способны при определенных условиях выбрасывать в окружающую среду свои лизосомы. При этом в одних случаях образуются небольшие выросты плазмалеммы, содержащие лизосомы, которые затем отрываются и переходят в среду. В других случаях наблюдается инвагинация плазмалеммы вглубь клетки и захват ею лизосом, распложенных далеко от поверхности клетки.
Процессы эндоцитоза и экзоцитоза осуществляется при участии связанной с плазмолеммой системы фибриллярных компонентов цитоплазмы.
Рецепторная функция плазмалеммы. Это одна из главных, универсальных для всех клеток, является рецепторная функция плазмалеммы. Она определяет взаимодействие клеток друг с другом и с внешней средой..
Все многообразие информационных межклеточных взаимодействий схематически можно представить как цепь последовательных реакций сигнал-рецептор-вторичный посредник-ответ (концепция сигнал-ответ). Передачу информации от клетки к клетке осуществляют сигнальные молекулы, которые вырабатываются в одних клетках и специфически влияют на другие, чувствительные к сигналу (клетки-мишени). Сигнальная молекула - первичный посредник связывается с находящимися на клетках-мишенях рецепторами, реагирующими только на определенные сигналы. Сигнальные молекулы - лиганды - подходят к своему рецептору как ключ к замку. Лигандами для мембранных рецепторов (рецепторов плазмалеммы) являются гидрофильные молекулы, пептидные гормоны, нейромедиаторы, цитокины, антитела, а для ядерных рецепторов - жирорастворимые молекулы, стероидные и тиреоидные гормоны, витамин Д В качестве рецепторов на поверхности клетки могут выступать белки мембраны или элементы гликокаликса - полисахариды и гликопротеиды. Считается, что чувствительные к отдельным веществам участки, разбросаны по поверхности клетки или собраны в небольшие зоны. Так, на поверхности прокариотических клеток и клеток животных имеется ограниченное число мест с которыми могут связываться вирусные частицы. Мембранные белки (переносчики и каналы) узнают, взаимодействуют и переносят лишь определенные вещества. Клеточные рецепторы участвуют в передаче сигналов с поверхности клетки внутрь ее. Разнообразие и специфичность наборов рецепторов на поверхности клеток ведет к созданию очень сложной системы маркеров, позволяющих отличать свои клетки от чужих. Сходные клетки взаимодействуют друг с другом, поверхности их могут слипаться (конъюгация у простейших, образование тканей у многоклеточных). Клетки не воспринимающие маркеры, а также отличающиеся набором детерминантных маркеров уничтожаются или отторгаются. При образовании комплекса рецептор-лиганд активируются трансмембранные белки: белок преобразователь, белок усилитель. В результате рецептор изменяет свою конформацию и взаимодействует с находящимся в клетке предшественником вторичного посредника - мессенджером. Мессенджерами могут быть ионизированный кальций, фосфолипаза С, аденилатциклаза, гуанилатциклаза. Под влиянием мессенджера происходит активация ферментов, участвующих в синтезе циклических монофосфатов - АМФ или ГМФ. Последние изменяют активность двух типов ферментов протеинкиназ в цитоплазме клетки, ведущих к фосфорилированию многочисленных внутриклеточных белков.
Наиболее распространено образование цАМФ, под действием которого усиливается секреция ряда гормонов - тироксина, кортизона, прогестерона, увеличивается распад гликогена в печени и мышцах, частота и сила сердечных сокращений, остеодеструкция, обратное всасывание воды в канальцах нефрона.
Активность аденилатциклазной системы очень велика - синтез цАМФ приводит к десяти тысячному усилению сигнала.
Под действием цГМФ увеличивается секреция инсулина поджелудочной железой, гистамина тучными клетками, серотонина тромбоцитами, сокращается гладкомышечная ткань.
Во многих случаях при образовании комплекса рецептор-лиганд происходит изменение мембранного потенциала, что в свою очередь приводит к изменению проницаемости плазмалеммы и метаболических процессов в клетке.
На плазматической мембране находятся специфические рецепторы, реагирующие на физические факторы. Так, у фотосинтезирующих бактерий на поверхности клетки располагаются хлорофиллы, реагирующие на свет. У светочувствительных животных в плазматической мембране находится целая система фогорецепторных белков-родопсинов, с помощью которых световой раздражитель трансформируется в химический сигнал, а затем электрический импульс.
межклеточная адгезия, подвижность клеток, образование цитоплазматических выростов (микроворсинки, стереоцилии, реснички, киноцилии).
Миофибрилла - немембранная сократительная органелла, состоит из упорядоченно упакованных тонких (актиновых), толстых (миозиновых) нитей и ассоциированных с ними вспомогательных белков, образующих актомиозиновый хемомеханический преобразователь и обеспечивающих сокращение миофибрилл в скелетных мышечных волокнах и сердечных мышечных клетках (кардиомиоцитах).
Аксонема - немембранная сократительная органелла - основной структурный элемент реснички и жгутика. Аксонема состоит из 9 периферических пар микротрубочек и двух расположенных центрально одиночных микротрубочек. Обладающий АТФазной активностью белок динеин - компонент тубулиндинеинового хемомеханического преобразователя - входит в состав ручек, связанных с периферическими микротрубочками. Матрицей для организации аксонемы служит базальное тельце - аналог центриоли.
Протеосома - функциональный макрокомплекс нелизосомных мультикаталитических протеиназ, широко распространённых в цитоплазме клеток эукариот. Протеосомы регулируют деградацию внутриклеточных белков, вовлечённых в различные клеточные процессы (размножение, рост, дифференцировка, функционирование), а т акже удаление повреждённых, окисленных и аберрантных белков.
Апоптосома - гептамерная колесоподобная структура- функциональный макрокомплекс, активирующий каспазы при апоптозе (регулируемой гибели клеток).
Включения образуются в результате жизнедеятельности клетки. Это могут быть пигментные включения (меланин), запасы питательных веществ и энергии (липиды, гликоген, желток), продукты распада (гемосидерин, липофусцин).
Плазматическая мембрана
Молекулярный состав
Все биологические мембраны имеют общие структурные особенности и свойства. Согласно жидкостно-мозаичной модели, предложенной в 1972 г. Николсоном и Сингером, плазматическая мембрана - жидкая динамическая система с мозаичным расположением белков и липидов. Согласно этой модели,
белковые молекулы плавают в жидком фосфолипидном бислое, образуя в нём своеобразную мозаику, но поскольку бислой обладает определённой текучестью, то и сам мозаичный узор не жестко фиксирован; белки могут менять в нем свое положение. Толщина плазматической мембраны - примерно 7,5 нм (рис. 2-2).
Основу мембраны составляет билипидный слой; оба липидных слоя образованы фосфолипидами. Фосфолипиды - триглицериды, у которых один остаток жирной кислоты замещён на остаток фосфорной кислоты. Участок молекулы, в котором находится остаток фосфорной кислоты, называют гидрофильной головкой; участок, в котором находятся остатки жирных кислот - гидрофобным хвостом. Жирные кислоты в составе гидрофобных хвостов бывают насыщенными и ненасыщенными. В молекулах ненасыщенных кислот имеются «изломы», что делает упаковку бислоя более рыхлой, а мембрану - более текучей. В мембране молекулы фосфолипидов расположены строго ориентировано в пространстве: гидрофобные концы молекул обращены друг к другу (от воды), а гидрофилные головки наружу (к воде). Липиды составляют до 45% массы мембран.
Холестерин имеет чрезвычайно важное значение не только как компонент биологических мембран; на основе холестерина происходит синтез стероидных гормонов - половых, глюкокортикоидов, минералкортикоидов. Холестерин участвует в формировании рафтов (плотиков) - дискретных мембранных доменов, богатых сфинголипидами и холестерином. Рафты представляют собой жидкостно-упорядоченную фазу (участок плотноупакованных липидов) и обладают отличной от плазмолеммы плотностью и точкой плавления, так что они могут «плавать» - перемещаться в плоскости жидкостно-неупорядоченной плазмолеммы для выполнения определённых функций.
Помимо липидов, в состав мембраны входят белки (в среднем до 60%). Они
определяют большинство специфических функций мембраны;
- периферические белки расположены на наружной или внутренней поверхности билипидного слоя;
- полуинтегральные белки частично погружены в липидный билипидный слой на различную глубину;
- трансмембранные, или интегральные белки пронизывают мембрану насквозь.
Углеводный компонент мембран (до 10%) представлен олигосахаридными или полисахаридными цепями, ковалентно связанными с молекулами белков
(гликопротеины) или липидов (гликолипиды). Цепи олигосахаридов выступают на наружной поверхности билипидного слоя и формируют поверхностную оболочку толщиной 50 нм - гликокаликс.
Функции плазматической мембраны
Основные функции плазмалеммы: трансмембранный транспорт веществ, эндоцитоз, экзоцитоз, межклеточные информационные взаимодействия.
Трансмембранный транспорт веществ . Транспорт веществ через плазматическую мембрану - это двустороннее движение веществ из цитоплазмы во внеклеточное пространство и обратно. Трансмембранный транспорт обеспечивает доставку в клетку питательных веществ, газообмен, выведение продуктов метаболизма. Перенос веществ через билипидный слой происходит путём диффузии (пассивная и облегчённая) и активного транспорта.
Эндоцитоз - поглощение (интернализация) клеткой воды, веществ, частиц и микроорганизмов. Эндоцитоз также происходит при перестройке или разрушении участков клеточной мембраны. К морфологически различаемым вариантам эндоцитоза относят пиноцитоз, фагоцитоз, опосредованный рецепторами эндоцитоз с образованием окаймлённых клатрином пузырьков и клатрин-независимый эндоцитоз с участием кавеол.
Экзоцитоз (секреция) - процесс, когда внутриклеточные секреторные везикулы (одномембранные пузырьки) сливаются с плазмолеммой, а их содержимое освобождается из клетки. При конститутивной (спонтанной) секреции слияние секреторных пузырьков происходит по мере их образования и накопления под плазмолеммой. Регулируемый экзоцитоз запускается с помощьюопределённого сигнала, чаще всего вследствие увеличения концентрации ионов кальция в цитозоле.
Межклеточные информационные взаимодействия. Клетка, воспринимая различные сигналы, реагирует на изменения окружающей её среды изменением режима функционирования. Плазматическая мембрана - место приложения физических (например, кванты света в фоторецепторах), химических (например, вкусовые и обонятельные молекулы, рН), механических (например, давление или растяжение в механорецепторах) раздражителей внешней среды и сигнальных молекул информационного характера из внутренней среды организма. Сигнальные молекулы (лиганды) (гормоны, цитокины, хемокины) специфически связываются с рецептором
Высокомолекулярным веществом, встроенным в плазмолемму. Клетка-мишень при помощи рецептора способна узнавать лиганд и отвечать изменением режима функционирования при связывании этого лиганда с его рецептором. Рецепторы гормонов стероидной природы (например, глюкокортикоидов, тестостерона, эстрогенов), производных тирозина и ретиноевой кислоты локализуются в цитозоле.
ПЛАЗМАТИЧЕСКАЯ МЕМБРАНА - (клеточная мембрана плазмалемма), биологическая мембрана, окружающая протоплазму растительных и животных клеток. Участвует в регуляции обмена веществ между клеткой и окружающей её средой.
Кле́точная мембра́на (также цитолемма, плазмалемма, или плазматическая мембрана) - эластическая молекулярная структура, состоящая из белков и липидов. Клеточная стенка, если таковая у клетки имеется (обычно есть у растительных клеток), покрывает клеточную мембрану. Клеточная мембрана представляет собой двойной слой (бислой) молекул класса липидов, большинство из которых представляет собой так называемые сложные липиды - фосфолипиды. Молекулы липидов имеют гидрофильную («головка») и гидрофобную («хвост») части. При образовании мембран гидрофобные участки молекул оказываются обращены внутрь, а гидрофильные - наружу.
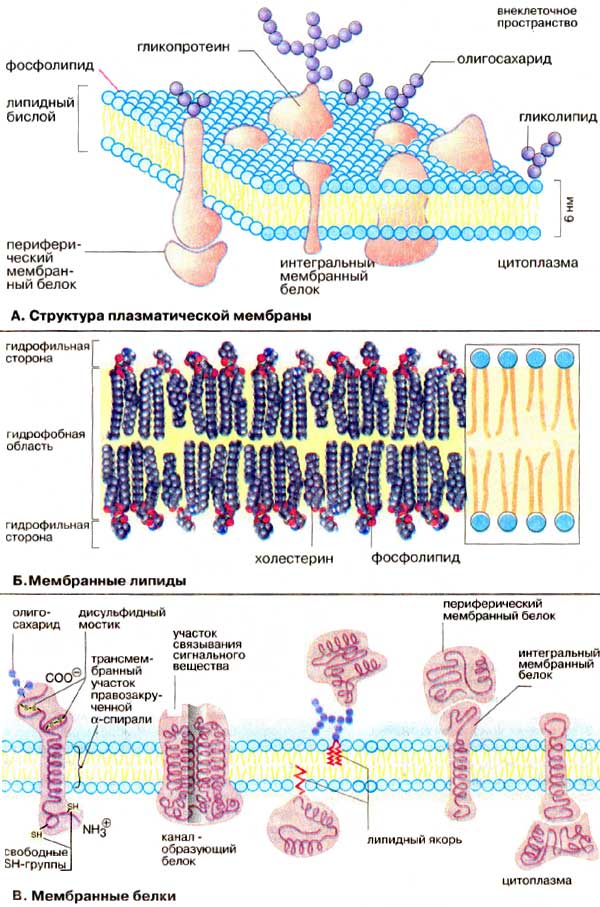
Структура мембраны клетки
Некоторое исключение составляют, пожалуй, археи, у которых мембраны образованы глицерином и терпеноидными спиртами. Некоторые белки являются точками контакта клеточной мембраны с цитоскелетом внутри клетки, и клеточной стенкой (если она есть) снаружи.
Смотреть что такое «плазматическая мембрана» в других словарях:
Эксперименты с искусственными билипидными пленками показали, что они обладают высоким поверхностным натяжением, гораздо большим, чем в клеточных мембранах. Дж. Робертсон сформулировал в 1960 году теорию унитарной биологической мембраны, в которой постулировалось трехслойное строение всех клеточных мембран.
Согласно этой модели белки в мембране не образуют сплошной слой на поверхности, а делятся на интегральные, полуинтегральные и периферические. Например, мембрана пероксисом защищает цитоплазму от опасных для клетки пероксидов. Избирательная проницаемость означает, что проницаемость мембраны для различных атомов или молекул зависит от их размеров, электрического заряда и химических свойств.
Вариантом этого механизма является облегчённая диффузия, при которой веществу помогает пройти через мембрану какая-либо специфическая молекула. Например, гормоны, циркулирующие в крови, действуют только на такие клетки-мишени, у которых есть соответствующие этим гормонам рецепторы. Нейромедиаторы (химические вещества, обеспечивающие проведение нервных импульсов) тоже связываются с особыми рецепторными белками клеток-мишеней.
С помощью маркеров клетки могут распознавать другие клетки и действовать согласованно с ними, например, при формировании органов и тканей. Мембраны состоят из липидов трёх классов: фосфолипиды, гликолипиды и холестерол.
Холестерол придаёт мембране жёсткость, занимая свободное пространство между гидрофобными хвостами липидов и не позволяя им изгибаться. Поэтому мембраны с малым содержанием холестерола более гибкие, а с большим - более жёсткие и хрупкие. Также холестерол служит «стопором», препятствующим перемещению полярных молекул из клетки и в клетку. Важную часть мембраны составляют белки, пронизывающие её и отвечающие за разнообразные свойства мембран.
Особенности обмена веществ в мембране
Рядом с белками находятся аннулярные липиды - они более упорядочены, менее подвижны, имеют в составе более насыщенные жирные кислоты и выделяются из мембраны вместе с белком. Без аннулярных липидов белки мембраны не работают. Избирательная проницаемость мембраны при пассивном транспорте обусловлена специальными каналами - интегральными белками. Они пронизывают мембрану насквозь, образовывая своего рода проход.
Относительно градиента концентрации молекулы этих элементов движутся в клетку и из неё. При раздражении каналы натриевых ионов раскрываются, и происходит резкое поступление в клетку ионовнатрия. Служит не только механическим барьером, но, главное, ограничивает свободный двусторонний поток в клетку и из нее низко- и высокомолекулярных веществ. Более того, плазмалемма выступает как структура, «узнающая» различные химические вещества и регулирующая избирательный транспорт этих веществ в клетку.
Механическая устойчивость плазматической мембраны определяется не только свойствами самой мембраны, но и свойствами прилежащих к ней гликокаликса и кортикального слоя цитоплазмы. Внешняя поверхность плазматической мембраны покрыта рыхлым волокнистым слоем вещества толщиной 3-4 нм - гликокаликсом.
В этом случае некоторые мембранные транспортные белки образуют молекулярные комплексы, каналы, через которые ионы проходят сквозь мембрану за счет простой диффузии. В других случаях специальные мембранные белки-переносчики избирательно связываются с тем или иным ионом и переносят его через мембрану.
ПЛАЗМАТИЧЕСКАЯ МЕМБРАНА - наружный слой цитоплазмы клетки более плотной консистенции. Заякоривающие соединения, или контакты, не только соединяют плазматические мембраны соседних клеток, но и связываются с фибриллярными элементами цитоскелета. Например, плазматические мембраны эпителиальных клеток кишечника содержат пищеварительные ферменты.
Какое строение имеет плазматическая мембрана? Каковы ее функции?
Основу структурной организации клетки составляют биологические мембраны. Плазматическая мембрана (плазмалемма) - это мембрана, окружающая цитоплазму живой клетки. Мембраны состоят из липидов и белков. Липиды (в основном фосфолипиды) образуют двойной слой, в котором гидрофобные «хвосты» молекул обращены внутрь мембраны, а гидрофильные - к её поверхностям. Молекулы белков могут располагаться на внешней и внутренней поверхности мембраны, могут частично погружаться в слой липидов или пронизывать её насквозь. Большая часть погруженных белков мембран - ферменты. Это жидкостно-мозаичная модель строения плазматической мембраны. Молекулы белка и липидов подвижны, что обеспечивает динамичность мембраны. В состав мембран входят также углеводы в виде гликолипидов и гликопротеинов (гликокаликс), располагающихся на внешней поверхности мембраны. Набор белков и углеводов на поверхности мембраны каждой клетки специфичен и является своеобразным указателем типа клеток.
Функции мембраны:
- Разделительная. Она заключается в образовании барьера между внутренним содержимым клетки и внешней средой.
- Обеспечение обмена веществ между цитоплазмой и внешней средой. В клетку поступают вода, ионы, неорганические и органические молекулы (транспортная функция). Во внешнюю среду выводятся продукты, образованные в клетке (секреторная функция).
- Транспортная. Транспорт через мембрану может проходить разными путями. Пассивный транспорт осуществляется без затрат энергии, путем простой диффузии, осмоса или облегченной диффузии с помощью белков- переносчиков. Активный транспорт - с помощью белков- переносчиков, и он требует затрат энергии (например, натрий-калиевый насос).
Крупные молекулы биополимеров попадают внутрь клетки в результате эндоцитоза. Его разделяют на фагоцитоз и пиноцитоз. Фагоцитоз - захват и поглощение клеткой крупных частиц. Явление впервые было описано И.И. Мечниковым. Сначала вещества прилипают к плазматической мембране, к специфическим белкам-рецепторам, затем мембрана прогибается, образуя углубление.
Образуется пищеварительная вакуоль. В ней перевариваются поступившие в клетку вещества. У человека и животных к фагоцитозу способны лейкоциты. Лейкоциты поглощают бактерии и другие твердые частицы.
Пиноцитоз - процесс захвата и поглощения капель жидкости с растворенными в ней веществами. Вещества прилипают к белкам мембраны (рецепторам), и капля раствора окружается мембраной, формируя вакуоль. Пиноцитоз и фагоцитоз происходят с затратой энергии АТФ.
- Секреторная. Секреция - выделение клеткой веществ, синтезированных в клетке, во внешнюю среду. Гормоны, полисахариды, белки, жировые капли, заключаются в пузырьки, ограниченные мембраной, и подходят к плазмалемме. Мембраны сливаются, и содержимое пузырька выводится в среду, окружающую клетку.
- Соединение клеток в ткани (за счет складчатых выростов).
- Рецепторная. В мембранах имеется большое число рецепторов - специальных белков, роль которых заключается в передаче сигналов извне внутрь клетки.





