Сложное строение человеческого мозга открывает тайны нашего поведения, объясняет законы мыслительной деятельности, протекание эмоций и чувств. Каждое полушарие головного мозга отвечает за свои специфические функции и задачи (например, известно, что правое отвечает за логику, а левое - за воображение и фантазию), но имеются также структуры, которые обеспечивают единую и согласованную работу всей центральной нервной системы. Одной из таких структур является ретикулярная формация.
Общие сведения
Ретикулярная формация - это участок стволового отдела мозга, представленный разветвленной сетью нервных клеток и ядер, соединяющих между собой различные отделы головного мозга. В отличие от других структур, - например, таламуса, гипоталамуса, мозжечка, - которые имеют некую цельную форму (ядра, железы), ретикулярная формация не представлена единым морфологическим образованием, а является «сетью» (от латинского reticulum — сеть) дендритов и аксонов, которые с разной степенью плотности проникают между отделами и структурами головного мозга, объединяя их между собой и обеспечивая их совместную деятельность.
Метафорически выражаясь: если наш мозг представить в виде некоего изделия, скажем, рубашки, то ретикулярная формация - это нити, которыми рубашка сшита. Ретикулярная формация пронизывает структуры продолговатого, среднего мозга и моста, имеет непосредственные связи с мозжечком, спинным мозгом, таламусом и опосредованные - с вышележащими отделами: гипоталамусом, зрительными ядрами и корой.
Как устроена
В состав ретикулярной формации входит огромное количество нейронов с разветвленными дендритами и длинными аксонами, за счет чего становится возможным передавать нервные импульсы в различные отделы головного и спинного мозга. При этом можно выделить две наиболее крупные группы нейронных скоплений:
- Ретикулотегментальное ядро, нейроны которого получают сигналы от вышележащих отделов ГМ (четверохолмие, таламус) и передают их далее в структуры мозжечка, регулируя тем самым некоторые жизненно важные двигательные функции: координация взгляда, движения глаз.
- Латеральное ядро, нейроны которого восходят от структур спинного мозга и вестибулярных ядер и обеспечивают информирование коры ГМ о положении тела в пространстве, участвуют в регуляции дыхания и сосудистой иннервации.
- Кроме того, в состав ретикулярной формации входят нейроны, которые принимают важное участие в работе центров терморегуляции, насыщения и голода.

Основные функции
Основное предназначение ретикулярной формации состоит в сенсорном анализе многочисленных сигналов, поступающих от различных отделов ГМ.
За счет тесных связей со спинным мозгом она также принимает самое активное участие в моторной регуляции, начиная от глотательного рефлекса и заканчивая сложными двигательными операциями. Кроме того, ретикулярная формация осуществляет активирующее воздействие на весь ГМ в целом, участвуя в регуляции циклов сна и бодрствования.
В общем виде функции ретикулярной формации сводятся к следующим:
- Регуляция скелетной мускулатуры (участвует в управлении движениями тела) и вегетативных функций (дыхание, чихание, кровообращение и т.п.).
- Контроль процессов сна и бодрствования (за счет оказания активирующего и тормозящего воздействия на кору больших полушарий).
- Активизирующая функция (проявляется в том, что ретикулярная формация обеспечивает постоянное тоническое возбуждение коры ГМ, за счет чего становится возможным поддержание внимания, сознания и протекание мыслительных процессов.)
- Обработка сигналов, поступающих из внешней и внутренней среды.
Отличительные особенности работы ретикулярной формации связаны, прежде всего, с определенными свойствами ее :
Нервные клетки, входящие в состав ретикулярной формации, обладают повышенной способностью к тоническому возбуждению. Это означает, что бо́льшая часть нейронов находятся в постоянном возбуждении и генерирует нервные импульсы, которые передаются в вышележащие отделы ГМ. Такая тоническая активность обусловлена рядом факторов:
- Проходимость большого количества сигналов через структуры ретикулярной формации. Приведем простую аналогию: представьте себе рояль или какой-либо другой струнный инструмент. Понятно, что когда мы напрямую касаемся струн, они начинают колебаться и издавать звук. То же самое происходит и с нервными клетками, когда к ним приходят сигналы от других нейронов.Однако представим далее, что мы не касаемся непосредственно струн инструмента, но, скажем, скачем рядом с ним, сильно стуча ногами о пол. Звука инструмента мы, может быть, и не услышим, но едва заметное колебание струн все же будет иметь место. То же самое происходит и с нейронами ретикулярной формации. Поскольку через нее постоянно проходят какие-то сигналы (как афферентные, так и эфферентные) от различных структур ЦНС, то это и создает постоянное тоническое возбуждение нейронов ретикулярной формации, за счет того, что она находится в эпицентре постоянного нервно-импульсного обмена.
- Повышенная чувствительность нейронов к химическим веществам (гормоны, медикаменты, психотропные вещества). Чашка кофе, выпитая утром, «включает» именно структуры ретикулярной формации и за счет длительного сохранения возбуждения в ее нейронах поддерживает нас в активном состоянии.
Нисходящее и восходящее влияние РФ
Как уже отмечалось, ретикулярная формация оказывает возбуждающее и тормозящее действие на различные отделы ГМ. При этом можно выделить два отдела, которые специализируются на передаче возбуждения в те или иные структуры мозга.
Нисходящий отдел: представлен вегетативными и двигательными центрами и оказывает нисходящее влияние на отделы спинного мозга. Соответствующие нейронные скопления регулируют деятельность дыхательного, сосудодвигательного, слюноотделительного центров, а также центров, отвечающих за построение простых и сложных двигательных реакций. Это указывает на решающую роль именно центральной нервной системы в регуляции даже элементарных безусловных рефлексов. Стимуляция нисходящего отдела приводит к затормаживанию спинальных центров и вызывает в естественной среде состояние глубоко сна (спать «без задних ног»). Тот же эффект можно вызвать и искусственным путем, например, вводя человека в состояние транса или наркоза.
Восходящий отдел: представлен нервными волокнами, соединяющими структуры ретикулярной формации с вышележащими отделами: таламусом, гипоталамусом, мозжечком и корой. Восходящее влияние оказывает стимулирующее действие на кортикальные структуры и обеспечивает активное состояние сознания. Восходящее влияние не прекращается, даже когда мы спим. Если бы наш мозг мог полностью «отключаться», то каждое пробуждение было бы сродни рождению: кто я? Где я? Как я сюда попал? Однако за счет работы ретикулярных структур у нас сохраняется возможность всегда возвращаться в то исходное состояние сознания, в котором мы находились до момента сна. Кроме того, и во время ночного отдыха у нас остается способность реагировать на некоторые жизненно важные раздражители, т.е. мы, как правило, не спим «мертвым сном» и можем проснуться, если рядом зашевелился и заплакал ребенок, что-то громко упало и т.п.
Проявление повреждения структур
Ретикулярная формация играет значимую роль в интегративной деятельности всего головного мозга. За счет выполнения функции ведущего проводника всех видов нервных импульсов во все отделы ЦНС, ретикулярная формация находится в постоянной работе. Чрезмерные умственные и эмоциональные перегрузки вредны для мозга в целом и для ретикулярной формации, в частности. К счастью, своевременный прием седативных препаратов может (за счет повышенной восприимчивости нейронов к химическому воздействию) быстро исправить ситуацию и нормализовать состояние.
Однако возможны и менее благоприятные исходы. Повреждение возможно в результате черепно-мозговых травм, онкологических заболеваний головного мозга, инфекционных поражений.
Основным проявлением неблагополучия является потеря сознания.
Нарушение восходящих связей обнаруживает себя в состоянии апатии, слабости, повышенной сонливости, двигательной растороможенности, нарушениях ночного сна. Нередки сопутствующие вегетативные расстройства.
Филогенетически очень древняя нейронная структура и хорошо развитый отдел ствола мозга рептилий. Сначала она представляла собой полисинаптический путь с медленным проведением, тесно связанный с обонятельной и лимбической областями. Прогрессирующее доминирование зрения и слуха над обонянием привело к смещению чувствительных и двигательных функций внутрь покрышки среднего мозга. Прямые спино-покрышечный и покрышечно-спинномозговой пути обходят ретикулярную формацию, которая отвечает, главным образом, за вегетативную регуляцию. У млекопитающих покрышка, в свою очередь, стала играть второстепенную роль в передаче возбуждения по очень быстро проводящим волокнам, соединяющим кору полушарий с периферическими двигательными и чувствительными нейронами.
В мозге человека ретикулярная формация сохраняет свою связь с лимбической системой и продолжает играть важную роль в вегетативной и рефлекторной регуляции.
Термин ретикулярная формация относят только к полисинаптической нейронной сети ствола мозга, несмотря на то, что сеть распространяется кпереди в таламус и гипоталамус и кзади в проприоспинальный тракт спинного мозга.
Общее строение показано на рисунке ниже. Срединная ретикулярная формация образована рядом ядер шва (греч.-nuclei raphe). Большая часть серотонинергических путей осевого отдела нервной системы начинается из ядер шва.
Ретикулярная формация (РФ).(А) Отделы. (Б) Группы аминергических и холинергических клеток.
Рядом расположена парамедианная ретикулярная формация. Этот отдел полностью состоит из крупноклеточных нейронов; в нижнем отделе моста и верхнем отделе продолговатого мозга (до уровня сращения ретикулярной формации с центральным ретикулярным ядром продолговатого мозга) можно также обнаружить гигантоклеточные нейроны.
Самым передним отделом считают латеральную мелкоклеточную ретикулярную формацию . Длинные дендриты мелкоклеточных нейронов образуют разветвления через определенные интервалы. Дендриты имеют преимущественно поперечное направление, а через промежутки между ними проходят длинные проводящие пути к таламусу. Латеральный отдел образован, главным образом, афферентными нейронами. К ним подходят волокна от всех чувствительных проводящих путей, включая органы чувств.
Обонятельные волокна проходят через медиальный пучок переднего мозга, расположенный рядом с гипоталамусом.
Зрительные проводящие пути проходят через верхний холмик.
Слуховые волокна подходят от верхнего ядра оливы.
Вестибулярные волокна подходят от медиального вестибулярного ядра.
Соматические чувствительные волокна проходят через спинно-ретикулярные тракты от спинномозгового и собственного (главного или главного мостового) ядер тройничного нерва.
Большая часть аксонов мелкоклеточных нейронов интенсивно разветвляется между дендритами нейронов парамедианной ретикулярной формации. Однако часть из них образует синапсы с ядрами черепных нервов и участвует в создании программ движений.
Парамедианная ретикулярная формация - преимущественно эфферентная система. Аксоны относительно длинные, некоторые поднимаются вверх, образуя синапсы с ретикулярной формацией ствола мозга или таламусом. От других отходят как восходящие, так и нисходящие ветви, образующие полисинаптическую сеть. К крупноклеточным нейронам подходят волокна от премоторной коры, которые дают начало ретикуло-спинномозговым путям моста и продолговатого мозга.

а) Аминергические нейроны ствола мозга . Рассеянные по ретикулярной формации группы аминергических (или моноаминергических) нейронов - нейроны, медиаторы которых образуются из ароматических аминокислот и обладают рядом влияний на клетку. Одна группа продуцирует нейромедиатор серотононин, три другие - катехоламины (дофамин, норадреналин и адреналин), одна группа - гистамин.
 Серотонинергические пути от срединного отдела ствола мозга (шва).
Серотонинергические пути от срединного отдела ствола мозга (шва).
Серотонинергические нейроны - наиболее часто встречаемые нейроны в любом отделе центральной нервной системы (ЦНС). К ним относят нейроны среднего мозга, волокна которых поднимаются к большим полушариям; нейроны моста, разветвляющиеся в стволе мозга и мозжечке; клетки продолговатого мозга, нисходящие в спинной мозг.
Все отделы серого вещества ЦНС пронизаны серотонин-секретирующими аксональными ветвями. Повышение серотонинергической активности используют в клинической практике для лечения такого распространенного заболевания, как большое депрессивное расстройство.

Дофаминергические нейроны среднего мозга представлены двумя группами. В месте соединения покрышки с ножками расположено черное вещество. Медиальнее него расположены вентральные ядра покрышки, от которых отходят мезокортикальные волокна к лобной доле и мезолимбические волокна, идущие непосредственно к прилежащему ядру.
Норадренергические (норэпинефринергические) нейроны немного менее многочисленные, чем серотонинергические. Около 90 % тел нейронов сконцентрировано в голубом пятне (locus ceruleus) в дне IV желудочка у верхнего конца моста. От голубого пятна начинаются пути во всех направлениях, что показано на рисунке ниже.
 Норадренергические пути от моста и продолгова того мозга.
Норадренергические пути от моста и продолгова того мозга.
Адреналин-секретирующие (эпинефрин-секретирующие) нейроны относительно немногочисленны и расположены преимущественно в ростральном/каудальном отделах продолговатого мозга. Одна часть волокон восходит к гипоталамусу, другая направляется книзу, образуя синапсы с преганглионарными симпатическими нейронами спинного мозга.
В больших полушариях ионная и электрическая активность аминергических нейронов значительно различается. Во-первых, для каждого амина существует более одного типа постсинаптических рецепторов. Во-вторых, некоторые аминергические нейроны высвобождают также белковые вещества, способные регулировать действие медиатора,- как правило, увеличивая его продолжительность. В-третьих, более крупные корковые нейроны получают множество возбуждающих и тормозных влияний от местных сетей с циркулирующим возбуждением, а также имеют множество различных типов рецепторов. Активация одного типа аминергического рецептора может привести к сильному или слабому эффекту в зависимости от исходного возбужденного состояния нейрона.
Наши знания о физиологии и фармакодинамике аминергических нейронов далеко не полные, однако их значение в широком разнообразии поведенческих функций не вызывает сомнений.
Часть поперечного среза через верхний отдел моста, показаны элементы ретикулярной формации.
Ретикулярная активирующая система (reticular activating system)
Ретикулярная формация (РФ) простирается от каудального отдела продолговатого мозга до рострального отдела промежуточного мозга. Ее нейроны образуют сетчатую структуру ствола мозга, в ячейках к-рой закреплены группы специфических ядер, снабжающих черепные нервы аксонами. Длинные восходящие и нисходящие тракты стволовой части головного мозга проходят через РФ и вблизи нее. Ее нейроны, в целом, имеют длинные аксоны, и одиночный нейрон РФ может иметь аксон, к-рый, разветвляясь, достигает ядер дорсального нервного пучка, спинного мозга и гипоталамуса. Вследствие такого широкого разветвления нейроны РФ способны оказывать глубокое воздействие на общий уровень активности головного и спинного мозга.
РФ не яв-ся ни функционально, ни структурно однородной. Ее можно разделить на группы ядер в соответствии с анат. и нейрохимическими критериями. Наиболее удобная анат. категоризация осуществляется по медиально-латеральному измерению. Ядра шва располагаются по срединной линии этой структуры. Крупноячеистая область находится в центре РФ, а мелкоячеистый отдел располагается в латеральной части РФ. Эти крупные медиальные/латеральные части можно подразделить на более мелкие отделы вдоль рострально-каудальной оси РФ. Напр., такое ядро, как п. gigantocellularis, яв-ся крупноячеистым ядром продолговатого мозга. Оно дает начало ретикулоспинальным аксонам, которые влияют на активность γ-мотонейронов. В такой структуре мозга, как мост, п. pontis caudalis и п. pontis oralis замещают п. gigantocellularis, тж давая начало ретикулоспинальным аксонам. Однако эти ядра, особенно рострально располагающееся п. pontis oralis, имеют многочисленные восходящие проекции и участвуют в регуляции кортикального возбуждения.
Нек-рые ретикулярные ядра могут тж определятся в соответствии с характером нейротрансмиттера. Обособленные группы нервных клеток, содержащих предполагаемый неиротрансмиттер норэпинефрин, были идентифицированы в РФ продолговатого мозга и моста. Наиболее широко изучено из них т. н. голубое пятно. Проекции из этого ядра распределяются по всему переднему мозгу и по мозжечку; как было установлено, они имеют отношение к REM-сну и к поддержанию интракраниальной самостимуляции. Допамин-содержащие нейроны обнаружены в покрышке среднего мозга; они тж имеют обширные проекции на передний мозг. Самая известная из этих допаминергических проекций берет начало в группе клеток, образующих черное вещество. Разрушение этого пути вызывает болезнь Паркинсона. Ядра шва дают начало серотонинергическим проекциям. Проекции переднего мозга из этих ядер включены в регуляцию медленноволнового сна, тогда как их спинномозговые проекции могут блокировать передачу болевых импульсов в головной мозг и производить изменения в функционировании автономной НС. Наконец, группы клеток в РФ среднего мозга дают начало восходящей холинергической проекции, к-рая участвует в регулировании неокортикального возбуждения, регистрируемого электрографическим способом.
См. также Головной мозг
М. Л. Вудрафф
Смотреть что такое "Ретикулярная активирующая система (reticular activating system)" в других словарях:
Совокупность нейронов и соединяющих их нервных волокон, расположенных в стволе мозга и образующих сеть. Ретикулярная формация связана со всеми органами чувств, двигательными и чувствительными областями коры большого мозга, таламусом и… … Медицинские термины
ФОРМАЦИЯ РЕТИКУЛЯРНАЯ, СИСТЕМА РЕТИКУЛЯРНАЯ АКТИВИРУЮЩАЯ - (reticular activating system) совокупность нейронов и соединяющих их нервных волокон, расположенных в стволе мозга и образующих сеть. Ретикулярная формация связана со всеми органами чувств, двигательными и чувствительными областями коры большого… … Толковый словарь по медицине
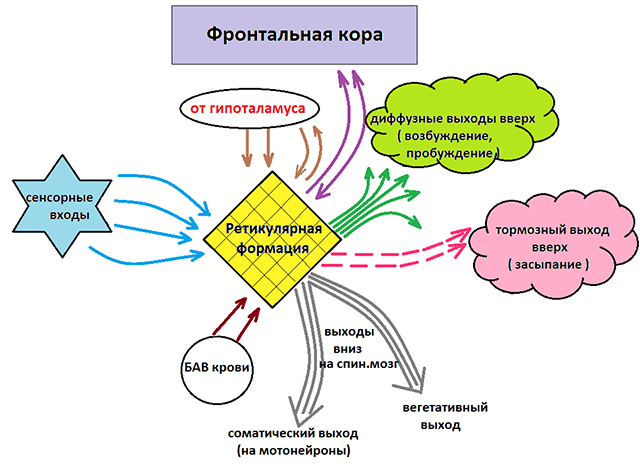
Ретикулярная формация - совокупность различных , расположенных на протяжении ствола мозга, оказывающих активирующее или тормозящее влияние на различные структуры центральной нервной системы, тем самым контролируя их рефлекторную деятельность.
Ретикулярная формация ствола мозга оказывает активирующее влияние на клетки и тормозное действие на мотонейроны спинного мозга. Посылая в спинной мозг к его двигательным нейронам тормозящие и возбуждающие импульсы ретикулярная формация участвует в регуляции тонуса скелетных мышц.
Ретикулярная формация поддерживает тонус вегетативных центров, интегрирует симпатические и парасимпатические влияния, передает модулирующее влияние от гипоталамуса и мозжечка к внутренним органам.
Функции ретикулярной формации
Соматодвигательный контроль (активация скелетной мускулатуры), может быть прямым через tr. reticulospinalis и непрямым через , оливы, бугорки четверохолмия, красное ядро, черное вещество, полосатое тело, ядра таламуса и даже соматомоторные зоны коры.
Соматочувствительный контроль , т.е. снижение уровней соматосенсорной информации — «медленная боль», модификация восприятия различных видов сенсорной чувствительности (слуха, зрения, вестибуляции, обоняния).
Висцеромоторный контроль состояния сердечно-сосудистой, дыхательной систем, активности гладкой мускулатуры различных внутренних органов.
Нейроэндокринная трансдукция через влияние на нейромедиаторы, центры гипоталамуса и далее гипофиз.
Биоритмы через связи с гипоталамусом и шишковидной железой.
Различные функциональные состояния организма (сон, пробуждение, состояние сознания, поведение) осуществляются посредством многочисленных связей ядер ретикулярной формации со всеми частями ЦНС.
Координация работы разных центров ствола мозга , обеспечивающих сложные висцеральные рефлекторные ответы (чихание, кашель, рвота, зевота, жевание, сосание, глотание и др.).
Строение ретикулярной формации
Ретикулярная формация образована совокупностью многочисленных нейронов , лежащих отдельно или сгруппированных в ядра (см. рис. 1 и 2). Ее структуры локализуются в центральных участках ствола, начиная с верхних сегментов шейного отдела спинного мозга до верхнего уровня ствола мозга, где они постепенно сливаются с ядерными группами . Ретикулярная формация занимает пространства между ядрами черепных нервов, другими ядрами и трактами, проходящими через ствол мозга.
Нейроны ретикулярной формации характеризуются большим разнообразием форм и размеров, но их общим признаком является то, что они образуют длинными дендритами и широко ветвящимися аксонами многочисленные синаптические контакты как между собой, так и с нейронами других ядер мозга. Эти ветвления формируют своеобразную сеть (ретикулум ), откуда произошло название — ретикулярная формация. У нейронов, формирующих ядра ретикулярной формации, имеются длинные аксоны , образующие проводящие пути к спинному мозгу, ядрам ствола мозга, и других областей головного мозга.

Рис. 1. Важнейшие структурные образования среднего мозга (поперечный срез)
К нейронам ретикулярной формации поступают многочисленные афферентные сигналы из различных структур ЦНС. Можно выделить несколько групп нейронов, к которым поступают эти сигналы. Это группа нейронов латерального ядра ретикулярной формации, расположенного в продолговатом мозге. Нейроны ядра получают афферентные сигналы от вставочных нейронов спинного мозга и входят в состав одного из непрямых спиномозжечковых путей. Кроме того, они получают сигналы от вестибулярных ядер и могут интегрировать информацию о состоянии активности вставочных нейронов, связанных с мотонейронами спинного мозга, и о положении тела и головы в пространстве.
Следующая группа — это нейроны ретикулотегментального ядра , расположенные на границе дорсального края моста. Они получают афферентные синаптические входы от нейронов претектальных ядер и верхних холмиков четверохолмия и посылают свои аксоны в структуры мозжечка, участвующие в контроле движений глаз.
Нейроны ретикулярной формации получают разнообразные сигналы через пути, связывающие их с корой головного мозга (кортикоретикулоспинальные пути), черной субстанцией, и .

Рис. 2. Расположение некоторых ядер в стволе мозга и гипоталамусе: 1 — паравентрикулярное; 2 — дорсомедиальное: 3 — преоптическое; 4 — супраоптическое; 5 — заднее
Кроме описанных афферентных путей в ретикулярную формацию поступают сигналы по аксонным коллатералям проводящих путей сенсорных систем. При этом на один и тот же могут конвергировать сигналы от разных рецепторов (тактильных, зрительных, слуховых, вестибулярных, болевых, температурных, проприорецепторов, рецепторов внутренних органов).
Из приведенного перечня основных афферентных связей ретикулярной формации с другими областями ЦНС видно, что состояние ее тонической нейронной активности определяется притоком практически всех типов сенсорных сигналов от чувствительных нейронов, а также сигналов от большинства структур ЦНС.
Классификация ретикулярной формации в зависимости от направлении волокон|
Отделы |
Характеристика |
|
Нисходящий отдел |
Вегетативные центры:
Двигательные центры:
|
|
Восходящий отдел |
Ретикулоталамические Ретикулогипоталамические Ретикуломозжечковые Ретикулокортикальные: активирующие; гипногенные |
Ядра ретикулярной формации и их функции
Долгое время считалось, что ретикулярная формация, строение которой характеризуется широкими межнейронными связями, интегрирует сигналы различной модальности, не выделяя при этом специфической информации. Однако становится все более очевидным, что ретикулярная формация является не только морфологически, но и функционально гетерогенной, хотя различия между функциями ее отдельных частей не столь очевидны, как это характерно для других областей мозга.
Действительно, многие нейронные группы ретикулярной формации формируют ее ядра (центры), выполняющие специфические функции. Это нейронные группы, формирующие сосудодвигательный центр продолговатого мозга (гигантоклеточное, парамедианное, латеральное, вентральное, каудальное ядра продолговатого мозга), дыхательный центр (гигантоклеточное, мелкоклеточное ядра продолговатого мозга, оральное и каудальное ядра моста), центры жевания и глотания (латеральное, парамедианное ядра продолговатого мозга), центры движений глаз (парамедианная часть моста, ростральная часть среднего мозга), центры регуляции тонуса мышц (ростральное ядро моста и каудальное — продолговатого мозга) и др.
Одной из важнейших неспецифических функций ретикулярной формации является регуляция общей нейронной активности коры и других структур ЦНС. В ретикулярной формации проводится оценка биологической значимости поступающих сенсорных сигналов, и в зависимости от результатов этой оценки она может активировать или тормозить через неспецифические или специфические нейронные группы таламуса нейронные процессы во всей коре головного мозга или в се отдельных зонах. Поэтому стволовая ретикулярная формация называется также активирующей системой ствола мозга. Благодаря этим свойствам ретикулярная формация может оказывать влияние на уровень общей активности коры, поддержание которой является важнейшим условием для сохранения сознания, состояния бодрствования, формирования направленности внимания.
Повышение активности ретикулярной формации (на общем высоком фоне) в отдельных сенсорных, ассоциативных областях коры обеспечивает возможность выделения и обработки специфической, наиболее важной в данный момент времени информации для организма и организации адекватных ответных поведенческих реакций. Обычно эти реакции, организуемые при участии ретикулярной формации ствола мозга, предваряются ориентационными движениями глаз, головы и тела в направлении источника сигнала, изменениями дыхания и кровообращения.
Активирующее влияние ретикулярной формации на кору и другие структуры ЦНС осуществляется по восходящим путям, идущим от гигантоклеточного, латерального и вентрального ретикулярных ядер продолговатого мозга, а также от ядер моста и среднего мозга. По этим путям потоки нервных импульсов проводятся к нейронам неспецифических ядер таламуса и после их обработки переключаются в таламических ядрах для последующей передачи к коре. Кроме того, от перечисленных ретикулярных ядер потоки сигналов проводятся к нейронам заднего гипоталамуса и базальных ганглиев.
Кроме регуляции нейронной активности высших отделов мозга ретикулярная формация может регулировать сенсорные функции. Это осуществляется путем влияния на проведение афферентных сигналов в нервные центры, на возбудимость нейронов нервных центров, а также на чувствительность рецепторов. Повышение активности ретикулярной формации сопровождается повышением активности нейронов симпатической нервной системы, иннервирующей органы чувств. В результате может повышаться острота зрения, слуха, тактильная чувствительность.
Наряду с восходящими активирующими и тормозящими влияниями на высшие отделы головного мозга ретикулярная формация принимает участие в регуляции движений , оказывая активирующие и тормозные воздействия на спинной мозг. На ее ядрах происходит переключение как восходящих путей, идущих от проприорецепторов и спинного мозга к головному мозгу, так и нисходящих двигательных путей от коры мозга, базальных ядер, мозжечка и красного ядра. Хотя восходящие нейронные пути, идущие из ретикулярной формации в таламус и кору, играют роль преимущественно в поддержании общего уровня активности коры больших полушарий мозга, но именно эта их функция важна для осуществления бодрствующей корой планирования, запуска, исполнения движений и контроля за их исполнением. Между восходящими и нисходящими через ретикулярную формацию путями имеется большое число коллатеральных связей, через которые они могут оказывать взаимное влияние. Существование такого тесного взаимодействия создает условия для взаимного воздействия области ретикулярной формации, влияющей через таламус на активность коры, планирующей и инициирующей движения, и области ретикулярной формации, влияющей на исполнительные нейронные механизмы спинного мозга. В ретикулярной формации имеются группы нейронов, которые посылают большинство аксонов в мозжечок, участвующий в регуляции и координации сложных движений.
По нисходящим ретикулоспинальным путям ретикулярная формация непосредственно воздействует на функции спинного мозга. Прямое влияние на его двигательные центры осуществляется по медиальному ретикулоспинальному тракту, идущему от ядер моста и активирующему преимущественно интер- и у-моторные нейроны разгибателей и тормозящему моторные нейроны мышц-сгибателей туловища и конечностей. По латеральному ретикулоспинальному тракту , начинающемуся от гигантоклеточного ядра продолговатого мозга, ретикулярная формация оказывает активирующее влияние на интер- и у-моторные нейроны мышц-сгибателей конечностей и тормозящее на нейроны мышц-разгибателей.
Из экспериментальных наблюдений на животных известно, что стимуляция более рострально расположенных нейронов ретикулярной формации на уровне продолговатого и среднего мозга, оказывает диффузное облегчающее влияние на спинальные рефлексы, а стимуляция нейронов каудальной части продолговатого мозга сопровождается торможением сиинальных рефлексов.
Активирующее и тормозное влияние ретикулярной формации на моторные центры спинного мозга может осуществляться через у-мотонейроны. При этом ретикулярные нейроны рострального участка ретикулярной формации активируют у-мотонейроны, которые своими аксонами иннервируют интрафузальные мышечные волокна, вызывают их сокращение, активируют рецепторы мышечных веретен. Поток сигналов от этих рецепторов активирует а-мотонейроны и вызывает сокращение соответствующей мышцы. Нейроны каудального участка ретикулярной формации тормозят активность у-мотонейронов спинного мозга и вызывают расслабление мышц. От баланса нейронной активности этих участков ретикулярной формации зависит распределение тонуса в больших мышечных группах. Поскольку этот баланс зависит от нисходящих влияний на ретикулярную формацию коры головного мозга, базальных ганглиев, гипоталамуса, мозжечка, то эти структуры мозга также могут через ретикулярную формацию и другие ядра ствола мозга воздействовать на распределение тонуса мышц и позу тела.
Широкое ветвление аксонов ретикулоспинальных путей в спинном мозге создает условия для влияния ретикулярной формации практически на все моторные нейроны и соответственно на состояние мышц различных частей тела. Такая особенность обеспечивает эффективное воздействие ретикулярной формации на рефлекторное распределение тонуса мышц, позу, ориентацию головы и тела в направлении действия внешних раздражителей и участие ретикулярной формации в осуществлении произвольных движений мышц проксимальных частей тела.
В центральной части ретикулярного гигантоклеточного ядра располагается участок, раздражение которого тормозит все двигательные рефлексы спинного мозга. Наличие такого торможения структур головного мозга на спинной мозг было открыто И.М. Сеченовым в опытах на лягушках. Суть опытов состояла в исследовании состояния рефлексов спинного мозга после перересечения ствола мозга на уровне промежуточного мозга и раздражении каудального участка разреза кристалликом поваренной соли. Оказалось, что двигательные спинальные рефлексы при раздражении не проявлялись или становились ослабленными и восстанавливались после устранения раздражения. Таким образом было впервые выявлено, что один нервный центр может тормозить активность другого. Это явление назвали центральным торможением.
Ретикулярная формация играет важную роль в регуляции не только соматических, но и вегетативных функций (ретикулярные ядра ствола мозга входят в структуру жизненно важных отделов дыхательного центра и центров регуляции кровообращения). Латеральная группа ретикулярных ядер моста и дорсолатеральное ядро покрышки формируют мочевыделительный центр моста. Аксоны нейронов ядер этого центра достигают преганглионарных нейронов крестцового отдела спинного мозга. Стимуляция нейронов этих ядер в мосту сопровождается сокращением мускулатуры стенки мочевого пузыря и мочевыделением.
В дорсолатеральном мосту расположено парабрахиальное ядро, на нейронах которого заканчиваются волокна сенсорных нейронов вкуса. Нейроны ядра, подобно нейронам голубоватого пятна и черной субстанции, содержат нейромеланин. Число таких нейронов в парабрахиальном ядре уменьшается при болезни Паркинсона. Нейроны парабрахиального ядра имеют связи с нейронами гипоталамуса, амигдалы, ядрами шва, одиночного тракта и другими ядрами ствола мозга. Предполагают, что парабрахиальные ядра имеют отношение к регуляции вегетативных функций и понижение их числа при паркинсонизме объясняет возникновение вегетативных нарушений при этом заболевании.
В опытах на животных было показано, что при раздражении некоторых локальных участков ретикулярных структур продолговатого мозга и моста можно вызвать торможение активности коры и сон. На ЭЭГ при этом возникают низкочастотные (1-4 Гц) волны. На основе описанных фактов считают, что важнейшими функциями восходящих влияний ретикулярной формации являются регуляция цикла сон — бодрствование и уровня сознания. Оказалось, что к формированию этих состояний имеют прямое отношение ряд ядер ретикулярной формации ствола мозга.
Так, с каждой стороны центрального шва моста располагаются парамедианные ретикулярные ядра, или ядра шва , содержащие серотонинергические нейроны. В каудальной части моста они включают нижнее центральное ядро, которое является продолжением ядра шва продолговатого мозга, а в ростральной части моста в состав ядер шва моста входит верхнее центральное ядро, называемое ядром Бехтерева, или срединным ядром шва.
В ростральной части моста на дорсальной стороне покрышки располагается группа ядер голубоватого пятна. В них имеется около 16 000-18 000 меланинсодержащих норадренергических нейронов, аксоны которых широко представлены в различных отделах ЦНС — гипоталамусе, гиппокампс, коре больших полушарий мозга, мозжечке и спинном мозге. Голубоватое пятно простирается в средний мозг, и его нейроны прослеживаются в ссром веществе околоводопроводного пространства. Число нейронов в ядрах голубоватого пятна уменьшается при паркинсонизме, болезни Альцгеймера и синдроме Дауна.
Как серотонинергические, так и норадреналинергические нейроны ретикулярной формации играют роль в контроле цикла сон — бодрствование. Подавление синтеза серотонина в ядрах шва ведет к развитию бессонницы. Предполагают, что серотонинергические нейроны являются частью нервной сети регуляции медленноволнового сна. При действии серотонина на нейроны голубоватого пятна возникает парадоксальный сон. Разрушение ядер голубоватого пятна у экспериментальных животных не ведет к развитию бессонницы, но вызывает на несколько недель исчезновение фазы парадоксального сна.





